Esplora le Cellule Staminali
In questa sezione forniamo un'introduzione alle cellule staminali, ai loro utilizzi, alla ricerca, alle schede informative, alle politiche in materia, alle news ed altri argomenti collegati.
Se non sai bene da dove iniziare, ti consigliamo di dare una occhiata alle schede informative.
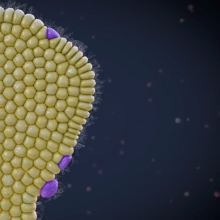






Speranze senza iperbole
• Risorsa di EuroStemCell
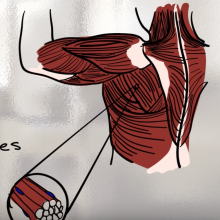
Da Dolly in poi
• Risorsa di EuroStemCell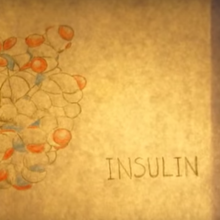
Coltura cellulare
• Risorsa di EuroStemCell
Conversazioni: etica, scienza e cellule staminali
• Risorsa di EuroStemCell
Cellule staminali - il futuro : introduzione alle cellule iPS
• Risorsa di EuroStemCell

Una storia di staminali
• Risorsa di EuroStemCell
Diabete: come possono essere di aiuto le cellule staminali?
• Risorsa di EuroStemCell
Le cellule staminali del cordone ombelicale: gli attuali utilizzi e le sfide future
• Risorsa di EuroStemCell
La ricerca sulle cellule staminali embrionali: un dilemma etico
• Risorsa di EuroStemCell
Rigenerazione: cosa significa e come avviene?
• Risorsa di EuroStemCell
Etica e riprogrammazione: questioni etiche dopo la scoperta delle cellule iPS
• Risorsa di EuroStemCell